Tuesday, 9 October 2012
The current state of my Ph.D. project
Enjoy my PREZI but don't get sick!
This presentation will be updated regularly and show the latest state of my PhD project on deep-sea isopods.
Cheers!
T
Monday, 16 May 2011
That's one small step for a man..
Today Buz Wilson and I were the first human beings to see the female genital opening of a deep-sea isopod crustacean belonging to the family Macrostylidae Hansen, 1916. Well, I reckon that's one small step for two men but one giant discovery for macrostylidologists. As I am to my knowledge the only person on this planet at the moment seriously specializing on this systematic group, I will have a drink on my own!
Wednesday, 4 May 2011
Towards a generalized species concept
An interesting paper about species concepts which I have just finished reading: http://onlinelibrary.wiley.com/doi/10.1111/j.1558-5646.2011.01231.x/full
I like Bernhard's approach very much. My enthusiasm is, however, a bit reserved regarding its practicability for my own studies. And I reckon it will be the same for many taxonomists / systematists working on other groups as well. It is the same with most other species concepts: We - the deep-sea isopod community - will never get to the point that we are in a position allowing us to apply the differential fitness species concept! How to infer the fitness effects of those few characters (be it morphology or molecules) which we are able to identify and use for systematics? Bernhard himself pointed at this problem in his final discussion and for us it remains the same procedure as before: We have to try our best to identify clusters based on morphological and molecular similarity, and maybe season it with some geographic evidence.
I like Bernhard's approach very much. My enthusiasm is, however, a bit reserved regarding its practicability for my own studies. And I reckon it will be the same for many taxonomists / systematists working on other groups as well. It is the same with most other species concepts: We - the deep-sea isopod community - will never get to the point that we are in a position allowing us to apply the differential fitness species concept! How to infer the fitness effects of those few characters (be it morphology or molecules) which we are able to identify and use for systematics? Bernhard himself pointed at this problem in his final discussion and for us it remains the same procedure as before: We have to try our best to identify clusters based on morphological and molecular similarity, and maybe season it with some geographic evidence.
Monday, 1 September 2008
Why taxonomy?
"The first step in wisdom is to know the things themselves; this notion consists in having the true idea of the object; objects are distinguished and known by their methodological calssification and appropriate naming; therefore Classification and Naming will be the foundation of our Science."
- LINNAEUS (1735), Quoted in Winston, 1999
I have nothing to add at the moment..
Cheers.. Torben
I have nothing to add at the moment..
Cheers.. Torben
Tuesday, 26 August 2008
Another enigmatic case.
Hi all! I start to feel a little bit like Christopher Taylor. I don’t know if this isn’t too much to be good for me and my progress in learning.
Again it is about crustacean parasites. Somebody ever heard about Tantulocarida? And again taxonomists had their problems with a taxon from the beginning of the 19th century until the 1980’s.? Not until then Tantulocarida where recognized as a discrete group of ectoparasites within the crustaceans. But as so many other “unlinkable” groups they can be found in the Maxillopoda today, following Schminke (2007). Unfortunately most of these articles I mention under “Reference” are not freely accessible or even not accessible trough the library online-system of my institute. But one of the leading scientists on Tantulocarida, Geoffrey Allan Boxshall, has written a summary on Answers.com.
These tiny crustaceans have crustacean hosts only and can be found sucking on Copepoda, Ostracoda, Cumacea, Tanaidacea and Isopoda in all oceans on earth.
The larva is called “Tantalus”. It is these larvae which have to find a new hosts. They attach to the hosts with a mouth disk and penetrate the host using an internal stylet which can be protruded out through the mouth opening. They suck haemolymph to feed or, as Boxshall points out, may have an “absorptive rootlet system” extending from the oral disc to penetrate the internal tissues of the host (Sounds a little bit like the rhizocephalan strategy).
It is known, that there are two enigmatic and separated reproduction strategies. A sexual lifecycle was inferred from the occurrence of a male and a female but in practice a reproduction as well as living adults were never seen.
Also, in sexual females the development from larva to adult was never observed. But it is assumed to happen like this: Soon after attaching to the host, the larva sheds its thorax. Instead, a huge sac develops on the Cephalon, out of which the adult female will hatch. (So basically it is a mixture of the developments of males and the second female type I will describe later.) Adult females have no appendages for swimming. The existence of a single big claw-like seta on the tip of each of the only two pairs of legs is interpreted as an organ to hold tight the male.
In males the larva does not shed its thorax but a strange metamorphosis takes place in a sack which develops behind the 5th or 6th segment of the larva. The sac is filled with a cluster of differentiated larval cells which reorganise to form an adult male. While the Cephalon is still attached to the host, an umbilical chord originating from the head supplies the male with nutrients from the host. By rupturing the sac wall the male is released. The adult male is able to swim, has 6 pairs of swimming legs, but it has no limb to hold the female. Males swim to find a female, it is assumed, and when he has found a receptive female, she grabs him. They have special sensory systems for that purpose, Aesthetascs, which possibly are derived antennae. Using their large penis, the male inseminate the female through a single ventral copulatory pore (Boxshall).
Inside a colossal carapace the female carries her eggs. Males and females – as it is usually implied – produce offspring in a sexual manner. It is assumed that the offspring is the Tantalus. Both, adult male and female have no anus so they do not seem to feed at all after hatching.
But there is also another type of female, which has a very different appearance compared to the sexual females. The so called “parthenogenetic female” is attached to the host permanently. The trunk of the larvae is most certainly shed after the attachment to the host, as it is in the sexual females. At least there is a scar found on the head. A sac-like new and unsegmented thorax develops behind the head. In this sac, which expands dramatically, tantalus-larvae are formed – without any molting - and released to infect new hosts, again without molting.
Whether the sexual or asexual phase is encountered and what controls it is unknown. Where is the link between these two phases? It is assumed that the asexual phase is encountered more frequently than the sexual phase, like in cladocerans or other Branchiopoda. And nobody knows what happens with the sexual female after its hatching. Does it hatch before or after a male has found it? When it hatches before: How does it avoid sinking to the sea floor?
Again it is about crustacean parasites. Somebody ever heard about Tantulocarida? And again taxonomists had their problems with a taxon from the beginning of the 19th century until the 1980’s.? Not until then Tantulocarida where recognized as a discrete group of ectoparasites within the crustaceans. But as so many other “unlinkable” groups they can be found in the Maxillopoda today, following Schminke (2007). Unfortunately most of these articles I mention under “Reference” are not freely accessible or even not accessible trough the library online-system of my institute. But one of the leading scientists on Tantulocarida, Geoffrey Allan Boxshall, has written a summary on Answers.com.
These tiny crustaceans have crustacean hosts only and can be found sucking on Copepoda, Ostracoda, Cumacea, Tanaidacea and Isopoda in all oceans on earth.
The larva is called “Tantalus”. It is these larvae which have to find a new hosts. They attach to the hosts with a mouth disk and penetrate the host using an internal stylet which can be protruded out through the mouth opening. They suck haemolymph to feed or, as Boxshall points out, may have an “absorptive rootlet system” extending from the oral disc to penetrate the internal tissues of the host (Sounds a little bit like the rhizocephalan strategy).
It is known, that there are two enigmatic and separated reproduction strategies. A sexual lifecycle was inferred from the occurrence of a male and a female but in practice a reproduction as well as living adults were never seen.
Also, in sexual females the development from larva to adult was never observed. But it is assumed to happen like this: Soon after attaching to the host, the larva sheds its thorax. Instead, a huge sac develops on the Cephalon, out of which the adult female will hatch. (So basically it is a mixture of the developments of males and the second female type I will describe later.) Adult females have no appendages for swimming. The existence of a single big claw-like seta on the tip of each of the only two pairs of legs is interpreted as an organ to hold tight the male.
In males the larva does not shed its thorax but a strange metamorphosis takes place in a sack which develops behind the 5th or 6th segment of the larva. The sac is filled with a cluster of differentiated larval cells which reorganise to form an adult male. While the Cephalon is still attached to the host, an umbilical chord originating from the head supplies the male with nutrients from the host. By rupturing the sac wall the male is released. The adult male is able to swim, has 6 pairs of swimming legs, but it has no limb to hold the female. Males swim to find a female, it is assumed, and when he has found a receptive female, she grabs him. They have special sensory systems for that purpose, Aesthetascs, which possibly are derived antennae. Using their large penis, the male inseminate the female through a single ventral copulatory pore (Boxshall).
Inside a colossal carapace the female carries her eggs. Males and females – as it is usually implied – produce offspring in a sexual manner. It is assumed that the offspring is the Tantalus. Both, adult male and female have no anus so they do not seem to feed at all after hatching.
But there is also another type of female, which has a very different appearance compared to the sexual females. The so called “parthenogenetic female” is attached to the host permanently. The trunk of the larvae is most certainly shed after the attachment to the host, as it is in the sexual females. At least there is a scar found on the head. A sac-like new and unsegmented thorax develops behind the head. In this sac, which expands dramatically, tantalus-larvae are formed – without any molting - and released to infect new hosts, again without molting.
Whether the sexual or asexual phase is encountered and what controls it is unknown. Where is the link between these two phases? It is assumed that the asexual phase is encountered more frequently than the sexual phase, like in cladocerans or other Branchiopoda. And nobody knows what happens with the sexual female after its hatching. Does it hatch before or after a male has found it? When it hatches before: How does it avoid sinking to the sea floor?
- Huys, R., G.A. Boxshall and R.J. Lincoln (1993); The Tantulocaridan Life Cycle: The Circle closed? Journal of Crustacean Biology, Volume 13, Issue 3, pp. 432–442.
- Boxshall, G.A. & R.J. Lincoln (1987); The Life Cycle of the Tantulocarida (Crustacea). Philosophical Transactions of the Roya Society of London. Series B, Biological Sciences, Volume 315, Issue 1173, pp. 267-303.
- Schminke,H.K. (2007); „Crustacea“ in Westheide, W. & R. Rieger’s „Spezielle Zoologie Teil 1. Einzeller und Wirbellose. Elsevier. Spektrum Akademischer Verlag 555-637.
Sunday, 24 August 2008
The Suckers, the Hijackers and the Limbless.
Crustaceans are THE group of organisms with most certainly the highest variability of body morphology and also a huge number if differences in habitat preferences, developmental modifications and life strategies. Many of these arthropods spend at least a part of their lifetime as parasites and this is what this post is about.
I am studying Westheide und Rieger's "Spezielle Zoologie" for several days now and I am amazingly surprised of the variability and number of taxa having parasitic lifestyles. Here’s about crustacean especially. But I am quite certain that other taxa will make me post as well.

{kind=link}
I am studying Westheide und Rieger's "Spezielle Zoologie" for several days now and I am amazingly surprised of the variability and number of taxa having parasitic lifestyles. Here’s about crustacean especially. But I am quite certain that other taxa will make me post as well.
Prof. H.K. Schminke from the University of Oldenburg, Germany wrote the chapter about Crustaceans. He keeps using a highly unresolved class called Maxillopoda to put in a couple of subtaxa which can so far not be linked to other Crustacean groups but are alltogether most likely no monophyletic group. Crustaceans most of them are. I am not joking. Included are coequally 9 subtaxa, namely the Mystacocarida, Copepoda, Branchiura (Fish Lice), Pentastomida (Tongue Worms), Ostracoda (Seed Shrimps), Tantulocarida, Facetotecta, Ascothoracida and Cirripedia (Barnacles and relatives). Other sources like the "Tree of Life web project" and some authors cited therein sum up Cirripedia, Facetotecta and Ascothoracida to a group called Thecostraca. But this doesn’t matter here. Fact is that some of these taxa have been mysteries until recently, at least for systematic zoologists.
The Facetotecta for example have until this year only been known because of their derived larvae, called y-nauplius (5 stages) and y-cypris (1 stage). These have been found worldwide and do not seem to be rare. Even though adult stages were never found so far, some larvae have been described as species and all the taxonomy and further systematic research on this group are based solely on the larvae. All described species belong to the genus Hansenocaris. The y-cypris has an undivided carapace and a “lattice organ”. The latter is most likely a chemo-sensory organ and links Facetotects to the barnacles and relatives. Its only extremities are the 1. antennae of which the 2nd segment may bear retractable claws. The labrum is long and also bears up to five hook-like claws covering the ventral cephalon. 6 pairs of thoracopods are made for swimming and the cypris has both, a single naupliar eye and a single compound eye. These appendages always were used to hypothesize a parasitic adult stage.
By reading this very interesting Blog by Christopher Taylor I was just pointed to a relatively new publication by Glenner et al. 2008 who found this previously unknown adult of a y-larva by induced metamorphosis. Well, it is no surprise that this adult stage of the so called Facetotecta was previously unknown despite the fact the its y-nauplius larva was known since the end of the 19th century. The authors point out that the animal is not capable of living on its own after metamorphosis and conclude it must be a parasite. But still nobody knows were to find the host. Isn’t that amazing? This kind of riddles is the reason for me to study biology and to do marine research, especially on crustaceans. Many mysteries remain unsolved!
More meshugga lifestyles: While barnacle-like cirripeds spend their adult life as sessile organisms on inorganic substrate (Thoracica) or buried in limestone or calcareous shells (Acrothoracica) filtering water, the Rhizocephala are a sistertaxon with a completely different lifestyle and another miracle of parasitism. Their relationship to other crustaceans and especially cirripeds was clarified by discovering their larval stages. But even to identify the adults as crustaceans is not possible on a morphological basis. Around
Female cypris larvae have to find the host – crabs in general. They penetrate soft parts of the Cuticula, usually joints on the ventral side of the pleon. These joints are not calcified and not sklerotized. When molting, the larvae grow into the host’s body. On the outside of the hosts cuticle a sack develops (Externa) as the posterior part and breeding organ of the parasite. Inside the host a root-system like anterior part (Interna) braids the internal organs of the host. Through this Interna the parasite feeds and also infers with the hormone system of the crab. The latter is sterilized and changes its body proportions (e.g. female-like pleons form in males).
Male rhizocephalans are dwarf-like and look different from the females even as larvae. After finding the mate, males swim to the Externa. Here they attach and leave their Cuticula - through the first Antennae which open at predetermined breaking points - as extremely small males. These are called Trichogons or dwarf males. The Trichogons become ameboid and penetrate the externa. By means of peristaltic movements they are moved to some receptacle, molt again and therewith close the entrance to keep away other males. The male is now implanting itself to the female body and starts producing sperm after some days. A female features two of these receptacles and can therefore incorporate two males. They will continue producing offspring together for a whole lifetime.
The male is nothing more but a sperm-producing amoeba living inside its mate. They invest 100% of “their” energy in sperm. The female keeps up to two males feeding them. 50% of her body is dedicated to rearing eggs and the other 50% are for manipulating the crab, which, male or female, form a broad Pleon as typical for females, not to take care of its own offspring but of the offspring of the parasites. Unbelievable!
In some other species the male larvae can inject their Trichogon through the cuticle of the female or even through the cuticle of the host when the female is located underneath. They accomplish that by means of their Antennae which they can use as an injection needle. You can find information about host specificity and an illustrated lifecycle of Sacculina carcini in a paper of Goddard et al. 2005. Well, it is especially about hosts in areas where it is found as an invader but still..
The Pentastomida are another subclass of “Maxillopoda” with not only obligatory parasitic stages but complete parasitic lifecycles. And this group was a riddle, too, as it was controversial for a long time, in which relationship to see them. They do not even have typical larvae to allocate them to crustaceans. And as the colloquial name “tongue worm” indicates their body plan is rather distinct from all other arthropods and even wormlike taxa among the invertebrates have not much in common with tongue worms.
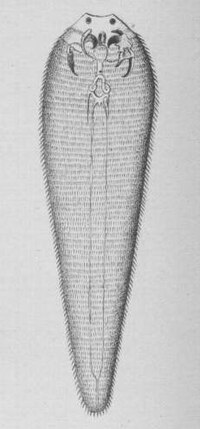{kind=link}
Tongue worms get up to 16cm in length and have a highly degenerated morphology. Adults live as ectoparasites in lungs, paranasal sinuses and trachea of terrestrial carnivore and omnivore vertebrates. Here they suck blood and mucus. 90% breed in reptiles, others in Canidae and Aves. The larvae live in many different terrestrial vertebrates of all groups and some Blattodea and Coleoptera. Linguatula serrata is common in dogs and may accidentally infect humans.
The latin name was caused by a misinterpretation of two pairs of openings next to the mouth opening comprising a retractable claw each. These claws may be homologous to Mandible and first Maxille in Arthropods. Except of additional frontal and apical papillae, which are interpreted as sensory system, there are no other appendages. There is no heart. Haemolymph is pumped by peristaltic movement of the whole body. There is no organ for excretion. The Body is annulated up to 230 times with two pairs of dorsoventral muscles for each annulus. This indicates a real metamerism. These are special developments which possibly are apomorphic characters for pentastomids. But these characters can be found nowhere else in crustaceans!
The Embryo has 4 pairs of appendage buds and a dorsal organ producing mucus. The four anterior buds develop to the frontal and apical papillae and the others make the claws. Anterior to the mouth the larva has a stiletto to penetrate the intestine wall of the host. Once it has found an alternate host or reinfected the original host, it moves through the body feeding and moulting several times. It develops to the adult stage gradually or another second maggot-like type of larvae may be interconnected. Alternate hosts normally are prey for the potential hosts.
Many works tried to find similarities between Pentastomida and any phylum of the animal kingdom or at least tried to eliminate possibilities. For example Karuppaswamy (1976) found β-Chitin in the cuticle of the Pentastomid Raillietiella gowrii using an X-ray technique and therefore delimited Pentastomulida from Arthropoda.
But finally, Lavrov et al. (2004) looked at the complete mtDNA sequence and gene arrangements and found a relation to Maxillopoda and especially Branchiura. This hypothesis remains to be tested. However, fossils from the Upper Cambrian identified as pentastomids (Walossek, D. and K. J. Müller, 1998) indicate an early branch dividing Pentastomida from the other Arthropods.
Whatsoever. Work remains to be done.
References:
- Walossek, D. and K. J. Müller (1998). "Cambrian "Orsten"-type arthropods and the phylogeny of Crustacea” in R. A. Fortey and R. H. Thomas: Arthropod Relationships. London: Chapman & Hall, 139–143.
- Goddard, J.H.R., M.E. Torchin, A.M. Kuris & K.D. Lafferty (2005); Host specificity of Sacculina carcini, a potential biological control agent of the introduced European green crab Carcinus maenas in
- Glenner, H., J.T. Høeg, M.J. Grygier & Y. Fujita (2008); Induced metamorphosis in crustacean y-larvae: Towards a solution to a 100-year-old riddle. BMC Biology
-
- Schminke,H.K. (2007); „Crustacea“ in Westheide, W. & R. Rieger’s „Spezielle Zoologie Teil 1. Einzeller und Wirbellose. Elsevier. Spektrum Akademischer Verlag 555-637.
-
Subscribe to:
Posts (Atom)